The origins of Wagyu cattle breeds
The Wagyu breed (和牛) has modest origins in Japan and the name translates in English to “Japanese cow”  but the excellence of eating quality has now become recognised and is in demand around the globe.
but the excellence of eating quality has now become recognised and is in demand around the globe.
Wagyu breeds were registered in Japan after a brief period of crossbreeding with native Japanese cattle from 1870s. “Improved Japanese Cattle” were recorded from 1919. The three modern breeds Japanese Black, Japanese Brown (Kochi strain)/Akaushi and Japanese Polled were fixed in 1944 by the Wagyu Registry Association so are distinct from the feral Kuchinoshima and Mishima that are native cattle. The regulations for Japanese Black classes were revised on 1st April, 1989 (Namikawa, K., 1992).
The Japanese Shorthorn Cattle Registry Association was established in 1957. Initially the breed was registered in 1945 in Iwate Prefecture as Tohoku Brown.
Origins of cattle from aurochs
Wild aurochs were present more than ten thousand years ago in different places in Eurasia and Africa. The aurochs were the large and powerful predecessors of domestic cattle then they became extinct in the 18th century. From the Bos primigenius genus they weighed between 300 and 800 kg, but were gradually driven out by people who used their domesticated relatives to exploit their habitat.
The aurochs were the large and powerful predecessors of domestic cattle then they became extinct in the 18th century. From the Bos primigenius genus they weighed between 300 and 800 kg, but were gradually driven out by people who used their domesticated relatives to exploit their habitat.
Taurine cattle (Bos taurus) were domesticated from Bos primigenius primigenius in the Upper Euphrates Valley in parts of modern day Syria, Iraq, Lebanon, Israel and Egypt during the Neolithic, more than 10,000 years ago. They are called ‘Near Eastern taurine’.
Approximately 1,500 years later a second domestication event took place in the Indus Valley in parts of India and Pakistan from Bos primigenius nomadicus, 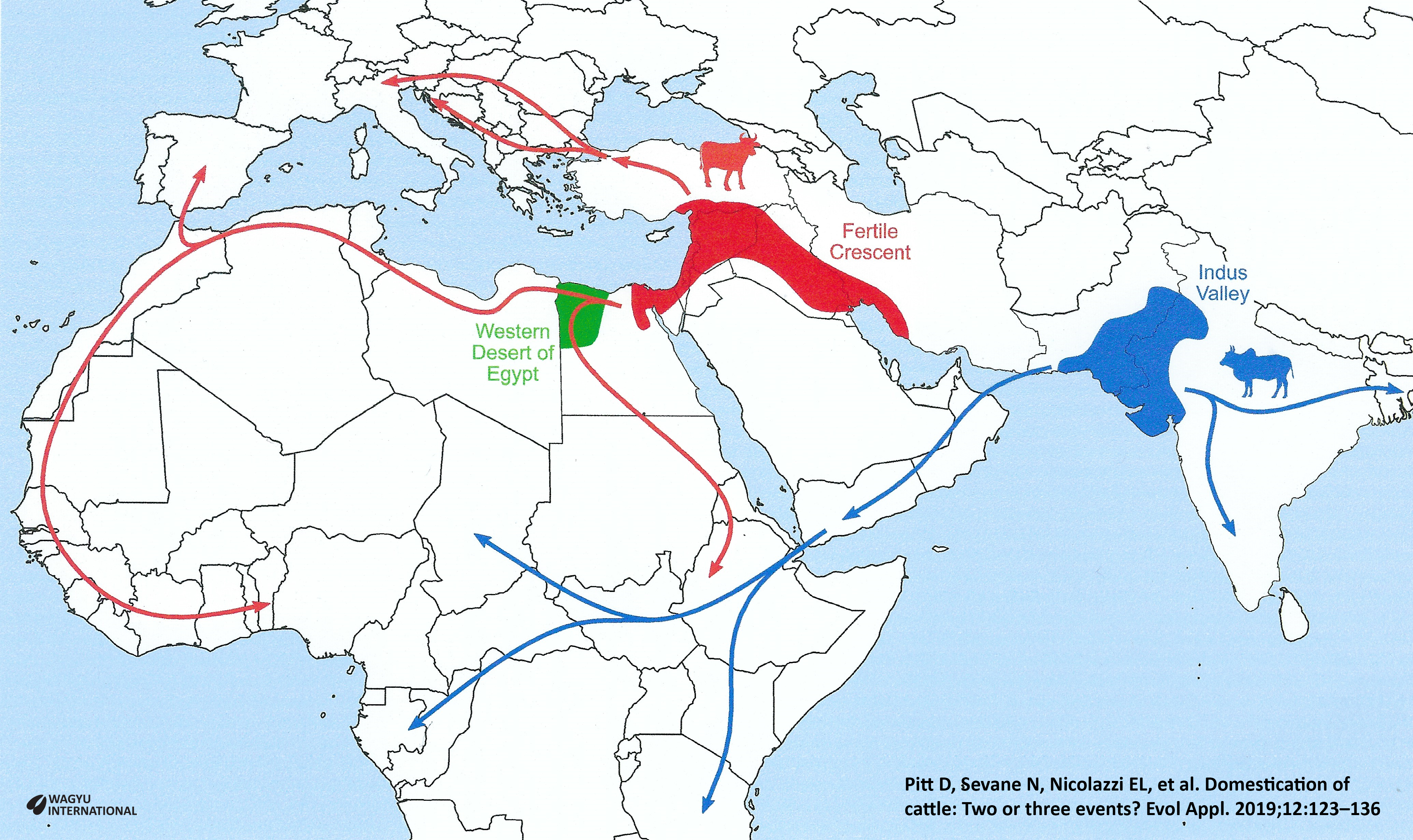 separating from the taurine branch to give rise to indicine cattle (Bos indicus) which are also known as Zebu cattle.
separating from the taurine branch to give rise to indicine cattle (Bos indicus) which are also known as Zebu cattle.
These are the two main domesticated events and a third is considered to have occurred in north-east Africa giving rise to the divergent African taurine cattle when there was admixture with indigenous African aurochs.
Iberian and Italian taurine resulted in the spread into Southern Europe across the Mediterranean Sea.
Migration and domestication of cattle in Asia
Katherine Brunson et alia posted pre-print on 10th August 2023 that cattle were sequenced after excavation from archaeological sites in eastern Mongolia. The 30,000 year old Paleolithic individual is the oldest data from an East Asia aurochs that was published to date. 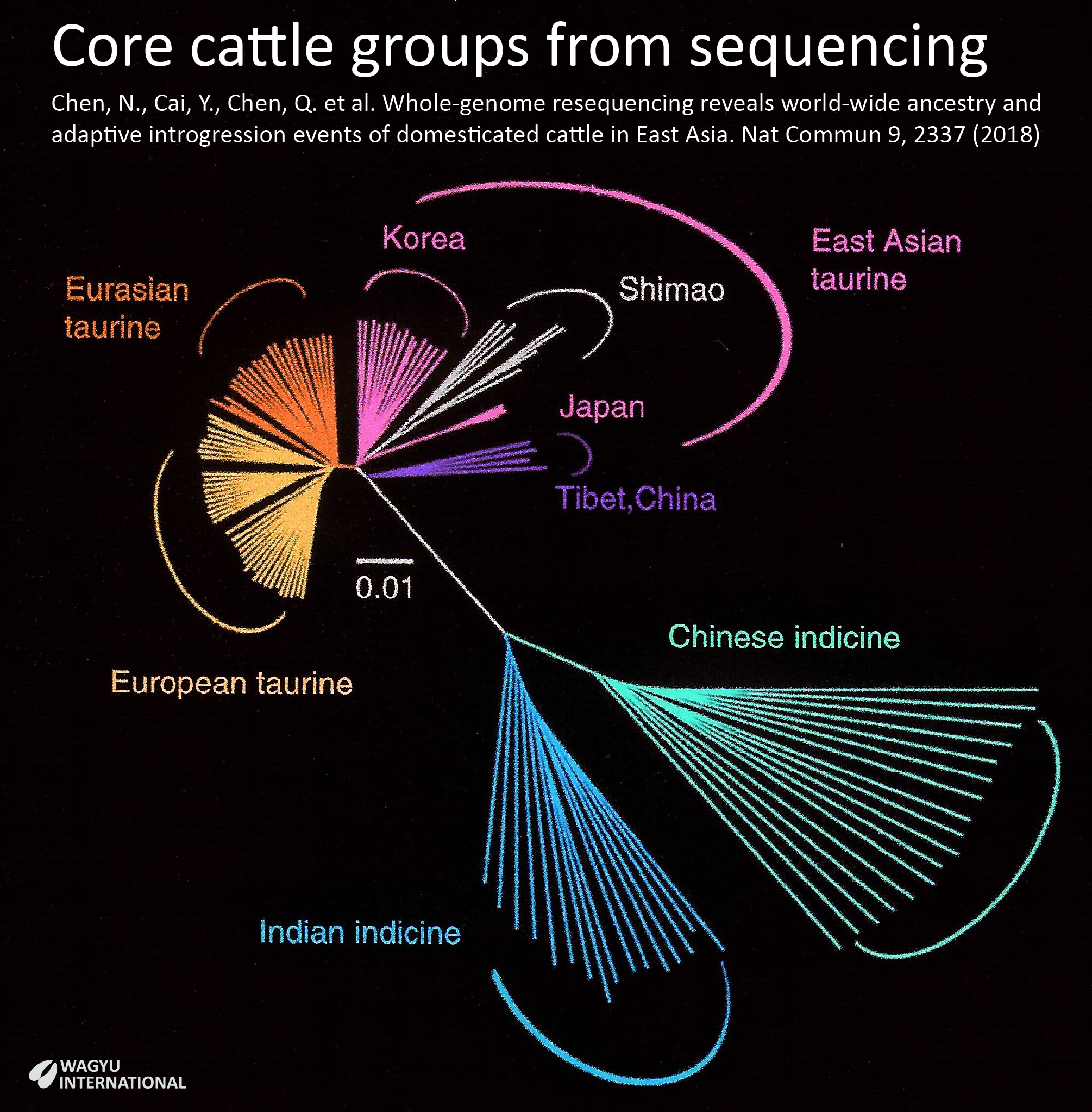
Early Neolithic samples were dated to between 8,000 to 5,500 years ago and these also pre-date domestication of cattle. There is growing archaeological evidence that people in Northeast Asia hunted and possibly managed wild aurochs from 12,000 BP until the adoption of domestic taurine cattle. On the Mongolian steppe, sedentary societies during the mid-Holocene relied heavily on aurochs, with increasingly sedentary groups continuing to specialize in big-game hunting.
Ancient taurine cattle from the site of Shimao in the Ordos region of China were dated to be 4,000 years and this is one the first sites in the Yellow River Valley Region to use domesticated cattle in large numbers.
However dogs were the first species to be domesticated in China. They were located in southern Hebei Province about 10,000 years before present (BP).
Pigs were the second species to be domesticated and were found in southern Henan about 9,000 BP.
Domestic sheep followed cattle and appeared in Gansu-Qinghai region about 5,600 to 5,000 years before present.
Domestic horses appear in eastern Gansu about 4,000–3,600 BP, and domestic chickens appear in north-eastern Henan about 3,300 BP.
Ningbo Chen et al., 2018, infers that two distinct B. taurus ancestries influenced East Asian cattle. The ancestral component labelled Eurasian taurine and the Y2a subhaplogroup were observed at the highest frequency in Northwest China and Central-South European cattle, whereas the Tibetan and Northeast Asian breeds shared the more narrowly distributed East Asian taurine ancestry component and the Y2b subhaplogroup. The earlier strand of ancestry was introduced several thousand years before. 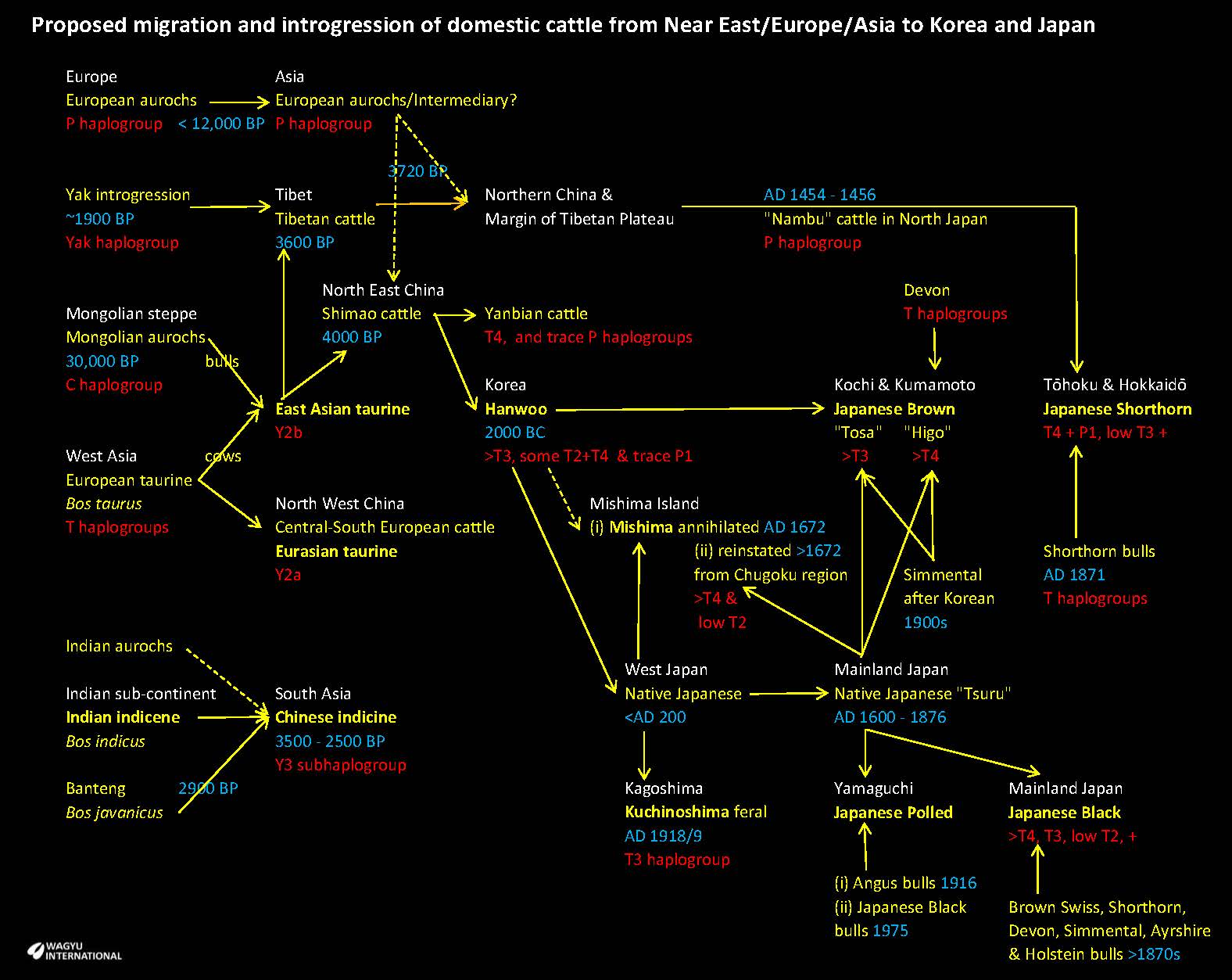
Later on, the East Asian taurine cattle became widespread all over northern China and margins of the Tibetan Plateau through broader expansion of millet agriculture around 3.6 thousand years ago. The expansion of early pastoralism, leading to west-to-east migration, may explain the similar component of Southern European and Mongolian cattle. In addition, the recent introgression of Mongolians into huge and extensive areas of China may have also led to gene flow across eastern Eurasia and partial replacement of East Asian taurine ancestry (and the Y2b lineage) in northern China.  The narrow distribution of pure East Asian taurine cattle in Tibetan Plateau and Northeast Asia might have been maintained by geographical barriers.
The narrow distribution of pure East Asian taurine cattle in Tibetan Plateau and Northeast Asia might have been maintained by geographical barriers.
Indicine cattle (Bos indicus), also called Zebu, may have dispersed from the Indian subcontinent to East Asia from 3,500 to 2,500 years before present. Chinese indicine cattle inherited around 2.93 % genome component from Banteng Bos javanicus ancestry at least 2.9 thousand years ago (Chen et al., 2018).
Previous population analysis based on genomic single nucleotide polymorphism (SNP) data revealed three major groups of cattle in the literature being Eurasian taurine, Asian indicine and African taurine
Chen et al., 2018 expanded this to five geographically distributed ancestral separations being European taurine, Eurasian taurine, East Asian taurine, Indian indicine and Chinese indicine.
| Major “core” groups/Separations & crosses | Region |
Modern breeds |
European taurine |
West European |
Hereford |
Eurasian taurine |
Central-South European |
Piedmontese |
North-West Chinese (Y2a) |
Kazakh Chaidamu |
|
East Asian taurine |
Tibetan (Y2b) |
|
North-East Asian (Y2b) |
Korean Hanwoo |
|
Indian indicine |
Indian (Y3b) |
Hariana |
Chinese indicine |
Chinese |
Leiqjiang |
Indian-Chinese indicine cross |
South China (Y3a) |
Dianzhong |
B. taurus and B. indicus hybridization |
Middle East |
|
Twenty years ago, Mannen et al., 2004 reported T4 is an East Asian haplogroup with a common, phylogenetically central haplotype that has not been observed in Africa, Europe or the Near East. The Mongolian and Japanese Black are composed of T, T2, T3 and T4 while the Korean network comprises T2, T3 and T4. T1 is dominant in Africa. In Japan, T4 predominates with T3 next most common. In Korea the T3 haplogroup is predominant and T4 is only represented twice (6.7%).
Aoi Noda et al., 2018 reported the high incidence of P haplogroups that were observed in Japanese Shorthorn. Using the mtDNA D-loop sequences, B. taurus mtDNA was categorized into five sub-haplogroups (T, T1, T2, T3 and T4) and B. indicus mtDNA into two sub-haplogroups (I1 and I2). The predomestic divergence between B. taurus and B. indicus was estimated to be 330,000 years.
In addition to the two major haplogroups T and I, five other haplogroups P, Q, R, C and E have been identified. Haplogroup Q has been detected in ancient domestic cattle and modern cattle in Eurasia and Africa, and it is closely related to haplogroup T. Haplogroup R has only been observed in modern Italian cattle and is phylogenetically distinct from haplogroups P, Q, and T. Haplogroup C was reported in ancient northeast Chinese cattle dated to 10,660 BP. Haplogroup E was identified in an aurochs (<6,000 BP) from Germany.
Haplogroup P was one of the major haplogroups in European aurochs but has not been detected in modern cattle in Europe. However, haplogroup P has only been identified in three modern Asian cattle, two Korean cattle and one Chinese cow using a dataset in excess of 3,000 haplotypes. Therefore, haplogroup P in modern cattle is considered to be a remnant of introgression from wild aurochs into the early domesticated cattle gene pool. The identification and analysis of these non-T haplogroups are useful tools for evaluating independent cattle domestication events and/or additional gene introgression from wild aurochs other than those in the Fertile Crescent and the Indus Valley.
Sequencing and origins of Wagyu breeds and Korean Hanwoo
It is considered that there were two major separate taurine migrations that reached Japan.
Yanbian cattle that are mainly found in North East China (Choi et al., 2016) are genetically close to Korean Hanwoo so they are a likely source for domestication in Korea. The ancestral component labelled Eurasian taurine had reached West Asia from the Near East. The East Asian taurine ancestry component with the Y2b subhaplogroup migrated to Tibetan and North east China. Taurine bones were found in North east China by about 5300 years ago and the Yellow River Valley by about 4500 years ago (Brunson et al., 2023).
Native cattle have been raised on the Korean peninsula since 2000 BC. In Chen's study of 2018, the neighbour-joining tree revealed the close affinity between Korean Hanwoo and native Japanese cattle. The divergence time was narrowed towards the end of the second century and the domestication of cattle to Japan coincides with the introduction of rice cultivation.
The migration of Bos taurus from ‘Near Eastern taurine’ by introgression through ‘East Asian taurine’ to Japan has been traced but the discovery of P haplogroups in Japanese Shorthorn resulted in a further review of the literature.
Transportation of horses and cattle to North Japan for military purposes is documented to have taken place between 1454 and 1456 AD. Hundreds to thousands of cattle and horses were imported from Mongolia and Siberia in 1454–1456 to the northern part of Japan.
The European aurochs migration east during the Pleistocene commenced 12,000 BP. Aurochs, bears and elks were depicted in rock paintings from the Neolithic to the Bronze Age in South Siberia. Incorporation of P haplogroup in Asian aurochs to P1a haplogroup via Kazakhstan/Russia-China/Mongolia-China Asian cattle was dated to 3,720 BP in north east China. The discovery of a high frequency of 45.9% mtDNA haplogroup P in Japanese Shorthorn cattle is considered to have originated from the importation from Mongolia and Siberia in 1454–1456 AD to the northern part of Japan. The native cattle in the Tohōku and Hokkaido regions of North Japan became known as “Nambu” cattle.
Shorthorn bulls were introduced in 1871 in several prefectures on northern parts of Honshu Island of Tohōku region during the Meiji Restoration. Ayrshire and Devon were also introduced and the Japanese Shorthorn breed became the fourth Wagyu breed in Japan. Japanese Shorthorn have been clustered closer with European breeds that Japanese Black, Japanese Brown (Kumamoto and Kochi that are both distinct), Korean Hanwoo and Mongolian through principal component analysis (Yonesaka et al., 2016).
The detection of the recessive mutation Factor 11 (F11) in both Black Wagyu and Red Wagyu is evidence that there is shared genomics between Japanese Black and “Higo” strain of Japanese Brown. Lim et al., 2016 did not detect Chediak–Higashi Syndrome (CHS), Spherocytosis, Claudin-16 (CL16) deficiency, nor Factor XI (F11) deficiency in Korean bulls so the F11 mutation took place in native cattle in Japan after migration from Korea.
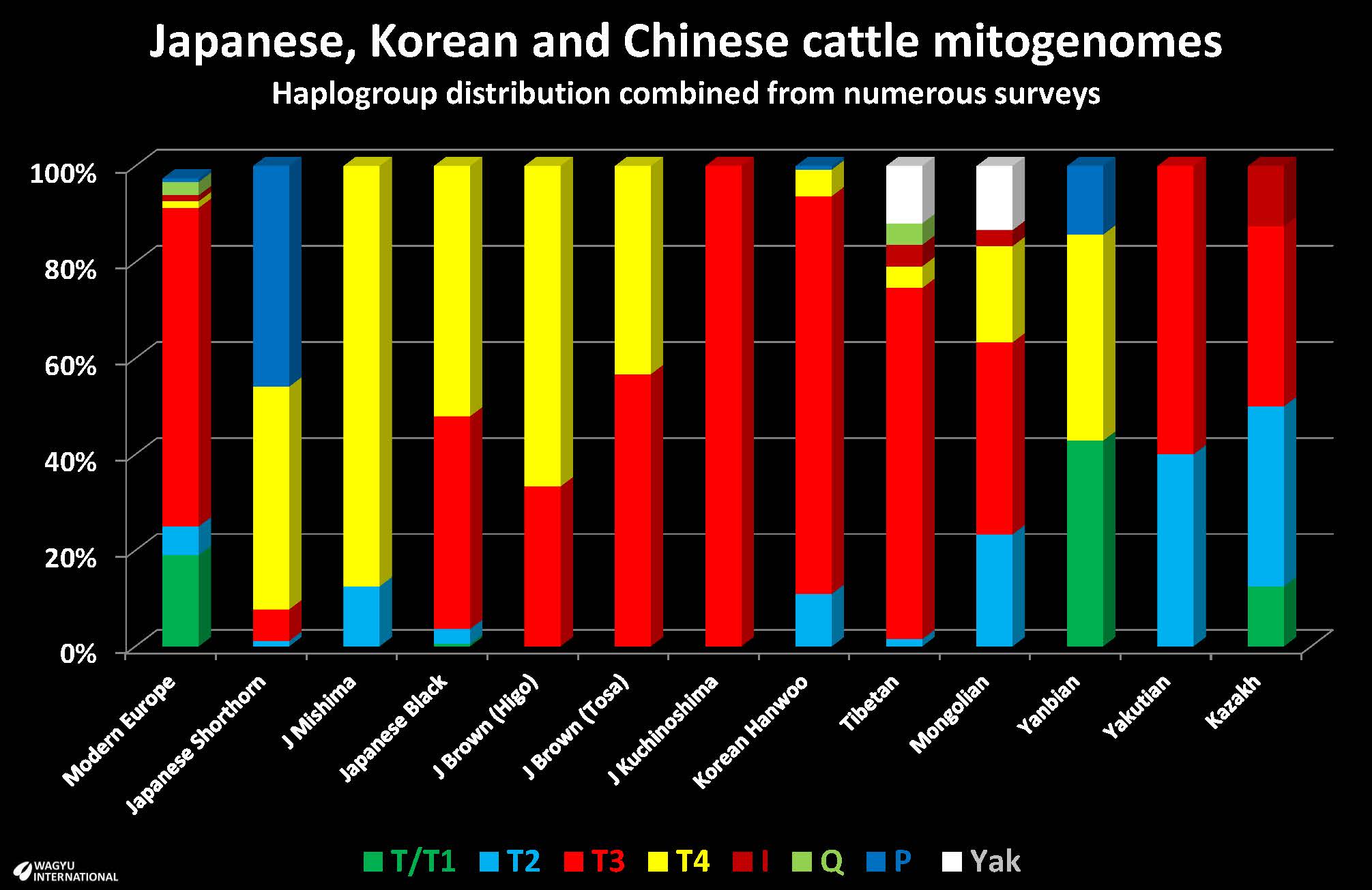
Minezawa reports genomic closeness (DA = 0.104) between Japanese Black and Japanese Brown despite considerable phenotypic differences that have resulted from selection after crossbreeding “improvements”. In Japan, “Higo” strain in Kumamoto, T4 haplogroup is predominant and this is the major haplogroup in Japanese Black and Simmental. The “Tosa” strain in Kochi, T3 is predominant and this is the major haplogroup in Korean Hanwoo.
Japanese Shorthorn is more distant from Japanese Black and this could reflect differences between "Nambu" and "Tsuru" ancestors, regionalisation and selection. DA is 0.196 between Japanese Black and Japanese Shorthorn, and 0.183 between Japanese Brown and Japanese Shorthorn.
History of beef cattle in Japan
Historians in the past wrote that pigs, cattle and chickens were introduced to Japan between the years 500 BC and 300 AD as domestic animals from the Asian continent by early immigrants. Chen et al., 2018 narrowed this down towards the end of the second century.
Cattle were used in mining, forestry, transportation and rice farming whilst horses were fundamentally used by the military. The selection pressure for draught capabilities resulted in this breed unintentionally evolving to produce the absolute ultimate in the beef tasting 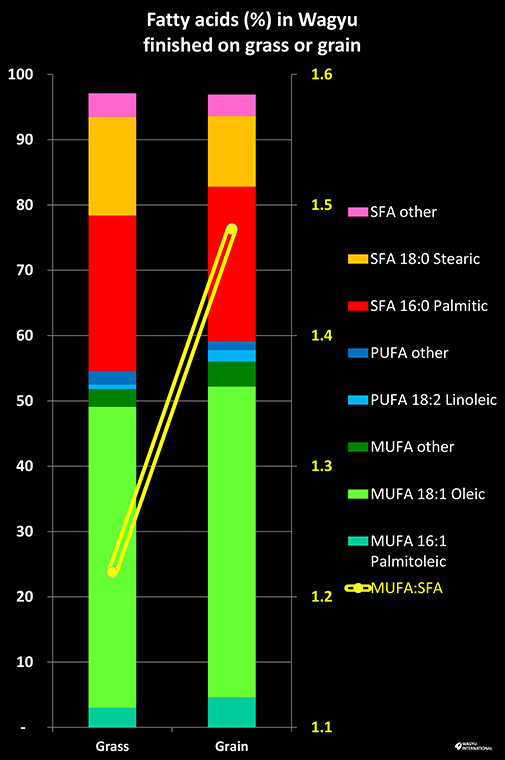 experience – for which they are now renowned around the world. Cattle that had stored fat within the muscles were able to work for longer and the most common lipids are monounsaturated fatty acids. Oleic acid (C18:1) is the most abundant fatty acid in marbled Wagyu from grass and grain feeding.
experience – for which they are now renowned around the world. Cattle that had stored fat within the muscles were able to work for longer and the most common lipids are monounsaturated fatty acids. Oleic acid (C18:1) is the most abundant fatty acid in marbled Wagyu from grass and grain feeding.
Buddhist leaders prohibited the eating of flesh - especially from four legged animals and there are records of banning the slaughtering of animals and chickens in AD 675. Slaughtering of cattle and horses was banned again in AD 742.
The native cattle became known as “Tsuru” and those in Okayama Prefecture were earning a premium in 1830 because of their physical prowess. Breeding was based on maternal traits as records were only kept of dams.
Lifting of the ban on meat consumption commenced during the Meiji era which started in 1867. Selection of cattle was principally on working performance and not for milk or meat production until mechanisation during the middle of the next century. Cattle were shipped from Kobe to Yokohama shortly after the port was opened there. The beef became popular with foreigners and it was called Kobe Beef. Even after the consumption of beef was permitted, cattle continued to be reared for draught purposes though some feeding commenced.
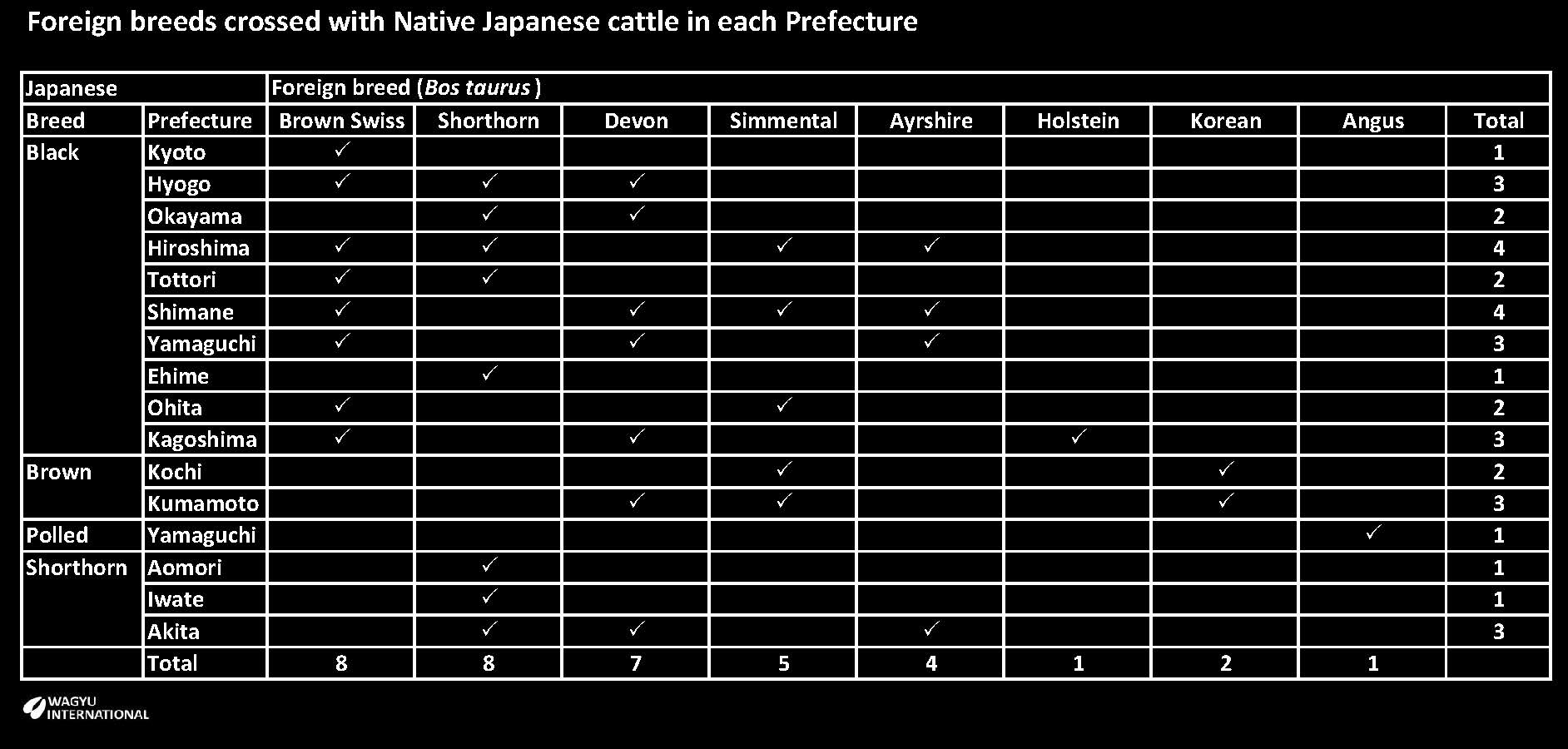
A decision was made to increase body size and milk production by introducing several dairy and beef breeds mainly from Europe. Initially Shorthorn and Devon bulls were imported in 1868. After a few years the government, prefectures and companies imported more breeds but there was neither planning nor coordination of the crossbreeding program. By 1887 about 2,600 foreign breeders had been introduced. The objectives of increasing body size and milking ability were quickly achieved so high prices were paid for crossbreds at the markets. Initial success was followed by failure when work performance and meat quality deteriorated so that most crossbreeding ceased within ten years after starting in each region. The foreign breeds are tabled by prefecture (from Minezawa, 2003 and Namikawa, 1992):
The government commenced registration and selection within the so called “Improved Japanese Cattle” in 1919. Type varied so extensively that the objectives were left to each prefectural organisation as an ongoing relic from the feudal age. Even though there was some integration gradually, the semen of active bulls seldom passed beyond prefectural borders. The three modern breeds Japanese Black, Japanese Brown (Tosa strain in Kochi prefecture) and Japanese Polled were established in 1944. 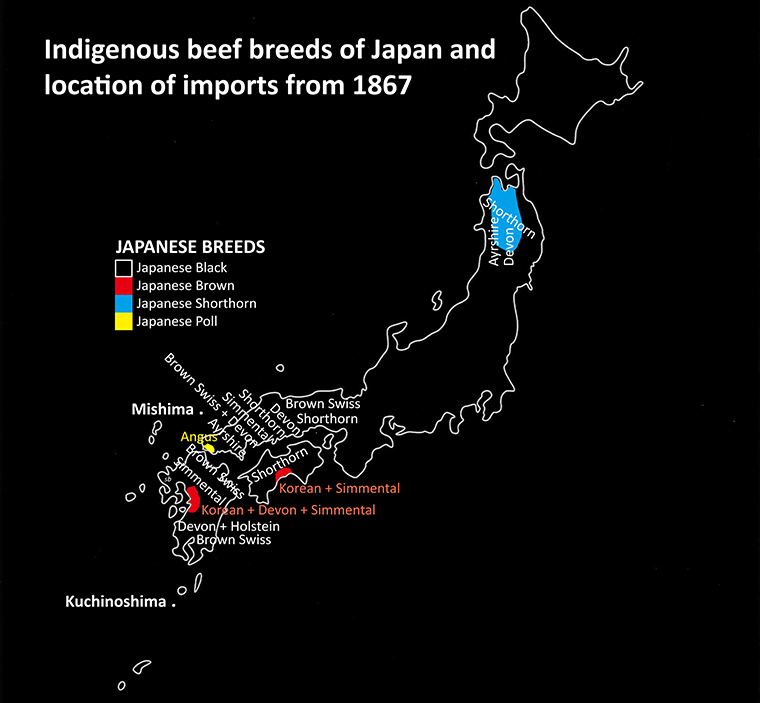 The fourth breed, Tohoku Brown was registered the following year in Iwate Prefecture but they were transferred in 1957 when the Japanese Shorthorn Cattle Registry Association was established.
The fourth breed, Tohoku Brown was registered the following year in Iwate Prefecture but they were transferred in 1957 when the Japanese Shorthorn Cattle Registry Association was established.
Breeding unions were started at village, town and county level. The committee determined which sires were used on each dam by visual assessment. Sasaki et al., 2005 outlined the criteria used for good visual judging: “Soft and elastic hide, fine and soft hair, fine textured horn and clean-cut face”. Some additional characteristics were reported by Namikawa 1992: “Black coat colour is predominant and black cattle with small white spot on udder or near anus, or a little amount of white hairs on black skin seemed to be preferred, rather than single black pattern... Quality of coat and skin were fine, and hair was curled...”
The following comments were noted on conformation: “Middle part of the body was plump with well stand (sic) ribs, but hind legs and neck were thin. Body shape was clean-cut, and shanks were fine with strong joint and tendon. Hooves were firm. Movement was smart.”
Since 1953 there has been a national show every five years and for the last twenty years it attracts a worldwide audience.
The development of fattening steers with some heifers replaced the former use of fattening cows after they had finished working. Until then, beef production had been a by-product from old cattle and co-incided with mechanization of agriculture from about 1950.
Results from progeny tests were recorded from 1968 when 111 were performance tested and 14 were progeny tested. This marked the end of visual phenotype selection and the beginning of some genetic improvement.
Until 1989, mature size and growth rate were the key measurements because of the close correlation with other measurements. 540 kg live weight and withers height of 129 cm was the objective at 35 months of age for mature cow size.
The widespread use of artificial insemination had generated feedback from field data of carcass results because more than 80% of slaughter steers were graded. 60,000 to 70,000 animals were registered each year so through best linear unbiased prediction (BLUP) analysis, predicted breeding values (PBVs) were issued. Each prefectural office managed the carcass data and PBVs from 1990. Variations between prefectures persisted and heritabilities of marbling ranged from 0.24 to 0.54 across 12 prefectures so PBVs so sires and dam could only be compared within each prefecture. BMS marble score PBV improved by 0.15 points annually in three prefectures and genetic trends from 1958 were reported by Sasaki et al., 2005.
This chart illustrates marbling in Japanese Black in Hyōgo and Oita and for Japanese Brown (Higo) in Kumamoto.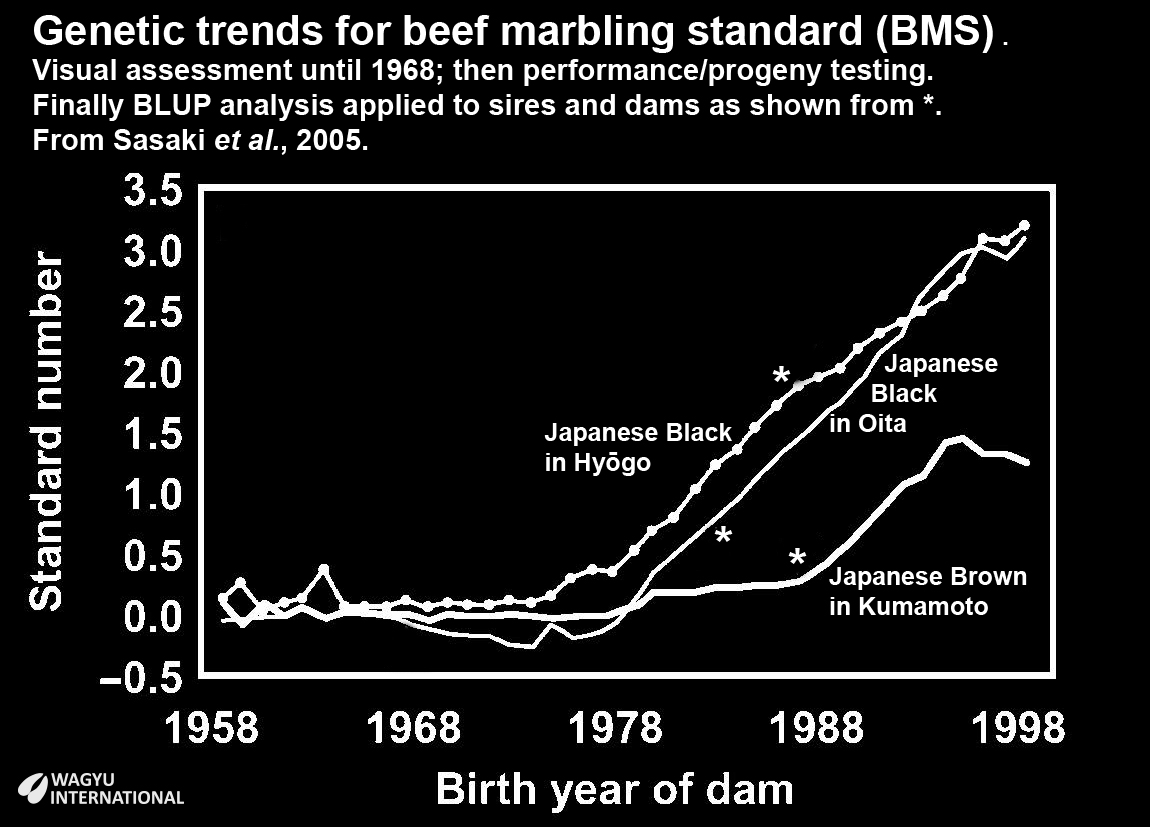
Selection within Japanese Brown was principally for size and this practice continued for at least another ten years after Beef Marble Score (BMS) was selected for in Japanese Black. Crossing with the imported European breed sires ceased over most of Japan after 1906 but there was a continuation of imports from Korea for the Tosa strain of Japanese Brown in Kochi. The Higo strain of Japanese Brown in Kumamoto was more influenced by Simmental and Devon breeds. Subsequently, persistent selection for size for eighty years has favoured the retention of Simmental genomics.
Standardized breeding values (SBV) were issued nationally so that breeding animals can be compared from different prefectures.
Despite improvements in breeding merit only becoming evident from 1970s, the dates that some of the key Japanese Black ancestors whose contributions were only recognised from that time put this history into perspective.
The ‘Nakadoi line’ is the benchmark for quality in Japan. The sire Nakadoi (中土井) was born in 1920 in Hyōgo. Seven generations later in 1939 in Hyōgo, the sire Tajiri (田 尻) was born. He is one of the most influential Japanese Black sires but he is also called Tajima in error in some pedigrees outside Japan. (Refer to Tafukudoi who was sired by Tajiri. Tajiri and his son have the same dam Fukue. Kenico ) The Anglophone name for the Tajiri family within Nakadoikei is “Tajima”. The ancestors of the Foundation sires that were exported that had the most influence back in Japan with year of birth are Kedaka 1959, Dai 7 Itozakura 1970, Dai 20 Hirashige 1974 and Yasufuku 1980.
References
Brunson, K., et al., 2016. New insights into the origins of oracle bone divination: Ancient DNA from Late Neolithic Chinese bovines. Journal of Archeological Science (74) 35-44.
Brunson, Katherine, Kelsey E., Susan Monge, Sloan Williams, David Peede, Davaakhuu Odsuren, Dashzeveg Bukhchuluun, Asa Carneron, Paul Szpak, Chunag Amartuvshin, William Honeychurch, Joshua Wright, Sarah Pleuger, Myagmar Erdene, Dashtseveg Tumen, Leland Rogers, Dorjpurev Khatanbaatar, Byambatseren Batdalai, Ganbaatar Galdan and Lisa Janz, 2023. Ancient Mongolian aurochs genomes reveal sustained introgression and management in East Asia. preprint version posted August 10, 2023.;https://doi.org/10.1101/2023.08.10.552443
Lim, Kyu-Sang, Park, B., Choi, T., Lim, D. and Y Cho, 2016. Carrier testing for autosomal recessive hereditary disorder in Korean proven bulls. J Bio Trans Res, (17) 4:85-90.
Mannen, H., Kohno, H., Nagata. Y., Tsuji, S., Bradley, D. G., Yeo, J. S., Nyamsamba, D., Zagdsuren, Y., Yokohama, M., Nomura, K. and T. Amano. 2004. Independent mitochondrial origin and historical genetic differentiation in North Eastern Asian cattle. Molecular Phytogenetics and Evolution. 32:539-544.
Mannen, H., Yonezawa, T., Murata, K., Noda, A., Kawaguchi, F., Sasazaki, S., Olivieri, A., Achilli, A and A. Torroni, 2020. Cattle mitogenome variation reveals a post-glacial expandsion of haplogroup P and an early incorporation into northeast Asian domestic herds. www.nature.com/scientificreports https://doi.org/10.1038/s41598-020-78040-8
McKay, S. D. , Schnabel, R. D. , Murdoch, B. M. , Matukumalli, L. K. , Aerts, J. , Coppieters, W. , Crews, D. , Dias Neto, E. , Gill, C. A. , Gao, C. , Mannen, H. , Wang, Z. , van Tassell, C. P. , Williams, J. L. , Taylor, J. F. and S. S. Moore. 2008. An assessment of population structure in eight breeds of cattle using a whole genome SNP panel. BMC Genetics 9:37.
Minezawa, M. 2003. Cattle genetic resources in Japan: One successful crossbreeding story and genetic diversity erosion. Cattle Genetic Resources in Japan. 4 (2):71-87.
Ministry of Agriculture, Forestry and Fisheries. Website.
Nagamine, Y., Nirasawa, K., Takahashi, H., Sasaki, O., Ishii, K, Minezawa, M., Oda, S., Visscher, P.M. and Furukawa, T. 2008. Estimation of the time of divergence between Japanese Mishima Island cattle and other cattle populations using microsatellite DNA markers. J. Hered. 99(2):202-207.
Namikawa, K. 1992. Breeding history of Japanese Beef cattle and preservation of genetic resources as economic farm animals. Wagyu. 2nd ed. Wagyu Registry Assoc., Kyoto, Japan.
Noda, A. Yonesaka, R., Sasazaki, S., H. Manneen, 2018. The mtDNA haplogroup P of modern Asian cattle: A genetic legacy of Asian aurochs? https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0190937
Nomura, T., Honda, T. and F. Mukai. 2001. Inbreding and effective population size of Japanese Black cattle. J. Anim. Sci. 79:366-370.
Ono, Kenichi, 1999. 100 Greatest Wagyu in Japan. Volume 1.
Oyama, K. 2011. Genetic variability of Wagyu cattle estimated by statistical approaches. An. Sci. J. 82:367-373.
Sasaki, Y., Miyake, T., and T. Dougo. 2006. Comparison of genetic gains per year for carcass traits among breding programs in the Japanese Brown and the Japanese Black cattle. J. Anim. Sci. 84:317-323.
Shojo, M., Okanishi, T., Anada, K., Oyama, K. and F. Mukai. 2006. Genetic analysis of calf market weight and carcass traits in Japanese Black cattle. J. Anim. Sci. 84:2617-2622.
Xia, X, K. Qu, G. Zhang, Y. Jia, Z. Ma, X. Zhao, Y. Huang, H. Chen, B. Huang, C. Lei, 2019. Comprehensive analysis of the mitochondrial DNA diversity in Chinese cattle. Wiley online library.
Yonesaka, R., Sasazaki, S., Yasue, H., Niwata, S., Inayousuke., Mukai, F and H Mannen, 2016. Genetic structure and relationship of 16 Asian and European cattle populations using DigiTag2 assay.
DISCLAIMER Wagyu International provides information that has been supplied by other parties and gives no warranty (express or implied) as to the data completeness, accuracy or fitness for a particular purpose.